A Temporary Convenience
A Critical Review of the Species Concept
William S. Abruzzi
(2015)

"I look at the term species as one arbitrarily given for the sake of convenience to a set of individuals closely resembling each other, and that it does not essentially differ from the term variety, which is given less distinct and more fluctuating forms."
--Charles Darwin (1859)
"Species are . . . a human contrivance, and the only value which the modern naturalist can attach to them, as such, is their temporary convenience as a means and vehicle of thinking and writing about the organic creation."
--
L. H. Bailey (1896)
* * * * *
Although species are widely viewed as genetic units defined by the ability of individual organisms to exchange hereditary material (DNA), this view of species does not accurately reflect the species concept as it is understood and applied in contemporary biological and ecological research. This paper critically examines the species concept and suggests that, given the arbitrariness of the definition of species and the inability to apply any species concept consistently and universally, species should be recognized not as a natural entity, but rather as a human construct with only limited validity and utility.
Most people view species as natural units defined by the ability of individual organisms to mate and produce fertile offspring, that is, possessing the ability to exchange and propagate hereditary material (DNA). This view of species does not accurately reflect the species concept as it is understood and applied in contemporary biological and ecological research. Contrary to the general understanding, interfertility is not the principal criterion used to determine whether two or more organisms are members of the same or different species. According to the late Ernst Mayr (1963:20), a proponent of the
Biological Species Concept (BSC) "species are more equivocally defined by their relation to non-conspecific populations ("isolation") than by the relation of conspecific individuals to each other. The decisive criterion is not the fertility of individuals but the reproductive isolation of populations." In other words, species status is determined not by whether two individuals can interbreed (interfertility), but rather by whether or not interbreeding normally occurs between members of two different populations in the wild (reproductive isolation).1
The distinction between interfertility and reproductive isolation is clearly illustrated by the case of the mallard (Anas platyrhynchos) and the pintail (Anas acuta), two of the most widely distributed freshwater ducks in North America (see Mayr 1963:90). Together, these two species comprise over 100 million individuals whose breeding ranges largely overlap. However, although they display complete interfertility in captivity, the two species are reproductively isolated in the wild, with the number of hybrids estimated at 1 in 1,000 (see Figure 1). Thus, "cross-fertility does not prove conspecificity." (Mayr 1963:91). Hubbs (1955:17) made essentially the same point regarding interbreeding among different species of fish.
"Merely placing a male of one species with a female of another species in an aquarium may lead to crossing. Thousands of such matings between swordtails and platies have been consummated, yet not a single hybrid between the two has ever been taken in nature, despite the fact that they often occur together and have been very extensively collected and very critically studied."
Figure 1

Mallard-Pintail Species Hybridization
Hybridization and Introgression
Indeed, interbreeding among members of different species (hybridization) is quite common (see Hubbs 1955; Bullini 1985, 1994;Arnold 1992, 1997). The tangelo and the nectarine are but two of the many domesticated plant species that have been created from the crossbreeding of two distinct natural species. The mule, the beefalo, and the liger (a cross between a male lion and a female tiger) are comparable examples of intentional animal species cross-fertilization. Grant and Grant (1992:195) indicate that over 1,500 different hybrid combinations of bird species alone have been bred in captivity.2
There has, of course, been a long history of domestic cross-fertilization of plant species, as well as the interbreeding of domesticated animals. However, considerable documentation exists of widespread hybridization in nature (see Stebbins 1959; Hubbs 1955; Short 1965, 1972; Bullini and Nascetti 1990; Wayne and Jenks 1991; Dowling and DeMarais 1993; Grant and Grant 1992, 1994; Roy et al. 1994; Dowling and Secor 1997; Arnold 1997). Documentation of species hybridization dates at least to Darwin (1859). Chapter 8 in The Origin of Species (titled "Hybridism") consists mostly of examples presented specifically to counter the claim that hybrid sterility serves as the defining criteria distinguishing species from varieties. In order to demonstrate this central point, Darwin presented numerous examples in which interbreeding between organisms classified as members of different varieties resulted in sterile hybrids together with a number of examples in which interbreeding between members of different species resulted in the production of fully fertile hybrids.
Humans have both inadvertently and intentionally introduced alien species into new territories, many of which have hybridized with native populations (Hubbs 1955; Abbott 1992; DeMarais et al. 1992; Dowling and DeMarais 1993; Smith 1992; Dowling and Secor 1997). Considerable hybridization also followed widespread changes in global climate that accompanied the retreat of the Pleistocene glaciers (Futuyma 1998: Chapter 16). Natural hybridization has been most common among plants (see Cronquist 1978), especially among such angiosperm (flowering plants) species as the iris and the sunflower (Arnold 1992:239-252).3 Grant (1981) has gone so far as to suggest that the majority of existing plant species originated from past hybridization (see also Ehrlich and Wilson 1991; Masterson 1994; Arnold 1997:24-25).4
While not as extensive as that among plant species, natural hybridization is far more common among animal species than is generally recognized. The mallard and pintail example presented above is by no means unique. Hybridization is especially common among birds and fish (Hubbs 1955; Short 1965, 1972; Smith 1992; Grant and Grant 1992, 1994; Dowling and DeMarais 1993; Dowling and Secor 1997). As Cronquist (1978:9-10) noted, "Although the amount of admissible hybridization among recognizable species of sunflowers may be greater than that among sparrows, the difference is only one of degree." Grant and Grant (1992:194, Table 1) list 895 out of 9,672 species of birds that hybridize, or 9.2% of the total. Mayr and Short (1970) claim that the level of hybridization among regularly hybridizing non-marine bird species averaged about 10%. In reality, however, the frequency of interspecific hybridization varies sharply among the different avian orders. While Short (1972) suggests that it was perhaps as high as 15% among some North American bird species, Arnold (1997:50-52) indicates that it is, in fact, as high as 19% among hummingbirds (Trochiliforms), 22% among grouse and partridges (Galliforms) and fully 42% among ducks and geese (Aseriforms). Similarly, according to Hubbs (1955), the frequency of hybridization among families of freshwater fish in North America ranged from as low as .3% for perch to as high as 17% among Pacific slope minnows.
Harrison (1993:5) defined hybridization as "the interbreeding of individuals from two populations, or groups of populations, which are distinguishable on the basis of one or more heritable characters." An important related concept, introgression, refers to "the permanent incorporation of genes from one set of differentiated populations into another, i.e. the incorporation of alien genes into a new, reproductively integrated population system" (Reisburg and Wendel 1993:71). While hybridization refers to gene mixing through interbreeding, introgression refers to any introduction of alien genetic material, including the gene transfers that occur: (1) between bacteriophage and their prey during symbiotic mycorrhizal associations (Van Valen 1976:237); (2) between insects and plants through viral exchanges (Anderson 1970; Zhdanov and Tikchonenko 1974); and (3) between carnivorans and primates (Todaro et al. 1974; Sherr and Todaro 1974; Benveniste and Todaro 1974; all referenced in Van Valen 1976:237). Smith (1992) discusses both the evolutionary and taxonomic consequences resulting from the extensive introgression in fish. The widespread occurrence of both hybridization and introgression in nature (see Anderson 1949; Arnold 1997), thus, severely undermines any notion that species represent discrete and integrated genetic systems.
In addition, it has been clearly documented that not all hybrids are sterile. According to Arnold (1997:3), "almost all cases of natural hybridization result in at least a few viable individuals with some measure of fertility."5 In other words, infertility is a matter of degree; as with any phenotypic characteristic, it varies quantitatively not qualitatively from one local population to another within the same species. At the same time, it is important to note when examining species hybridization that not all offspring of same-species matings are viable or fertile either. As Cronquist (1978:10) points out, "Self-sterile plants are usually also sterile with a certain fraction of the population to which they belong. Thus the inability of individuals a and b to cross with each other does not necessarily prevent both from being interfertile with individuals c, d, e, f, and g."
Furthermore, while some researchers, most notably Mayr (1942, 1963, 1996),6 have rejected the evolutionary significance of hybridization, others have argued that hybridization is not only widespread, but has, in fact, been a significant factor in both species diversification and the evolution of higher taxa (Grant and Grant 1992, 1994; Harlen and de Wet 1963; Dowling and Secor 1997). Indeed, the very finches studied by Darwin on the Galapagos Islands provide an important example of the evolutionary significance of hybridization. Grant and Grant (1992, 1994) found that in terms of survival, clutch size and hatching success, hybrid species of finches on Daphne Major Island in the Galapagos had a higher fitness than non-hybrids. Nagle and Mettler (1969) made a similar observation among various populations of Drosophila; one of their hybrid populations achieved a higher fitness than all of the other hybrids, as well as that of both parental species. Similarly, Hubbs (1955) documents extensive hybridization between different sunfish species, which he attributes to heterosis (hybrid vigor). Hybrids between the green sunfish and the bluegill developed by Hubbs grew faster than either parental species. They were also larger and more vigorous in taking food and ranked higher in the social hierarchy (nip order).
The evolutionary significance of hybridization is based on the fact that while hybrids might be less well adapted under one set of circumstances, they may be better adapted under a different set of conditions. Hubbs (1955) found significant differences in the rate of hybridization between marine fishes and northern fresh water fish, as well as between fish species inhabiting coastal streams vs. those in the interior. Whereas marine fish hybrids were "excessively rare," Hubbs (1955:2) discovered that 95% of the sunfish he examined in one stream near Ann Arbor were hybrids between the green sunfish and pumpkinseeds. Hubbs attributed the more frequent hybridization among freshwater fish in large part to the greater opportunity for interbreeding between different species created by the changing nature of interior lakes and streams as compared to the greater stability of oceans. In general, hybrids are found in intermediate habitats where more specialized parental species are not as adaptable. Similarly, "environmental heterogeneity and instability are conducive to the formation of hybrid species" (Dowling and Secor 1997:606).
Durden (1969) has argued that while individual traits might change through gradual evolution, species change is more likely to occur in leaps, representing "jumps" from the optimal occupancy of one niche to the optimal occupancy of another distinct but similar niche. Grant and Grant (1994:313) similarly argue that the ability of populations to respond to environmental changes and to evolve in new directions is
". . . constrained by the architecture of development under the influence of genes; therefore, the assembly of new gene combinations and the creation of genetic novelty by hybridization has the potential of relaxing the constraints, of permitting altered developmental programs, and hence altered genetic correlations between traits."7
Hybridization and introgression increase genetic diversity more quickly than can be achieved through mutations (Grant and Grant 1994:312-313; Dowling and Secor 1997:610), allowing organisms greater flexibility in adapting to changing environmental conditions and opening up opportunities for new evolutionary lines. This feature is of particular evolutionary significance, given the important connection between environmental change and the origin of new species and higher taxa, such as occurred following the recent retreat of the glaciers.8 Hubbs (1955:6) has shown how this process works. Whereas cutthroat and rainbow trout live side by side with little or no hybridization in coastal streams, the introduction of rainbow into interior bodies of water inhabited by cutthroat in the absence of rainbow led to extensive hybridization and frequently to the complete elimination of cutthroat.
An assessment of chromosomal and genetic evidence suggests that speciation through hybridization may have contributed to the origin of several insect species, including walking sticks, grasshoppers, black flies and cucurlionid beetles (Bullini and Nascetti 1990; see also Boxhorn 1995:7). Similarly, the recent analysis of mitochondrial DNA (mtDNA) among the Florida panther and 10 other North American and South American panther subspecies also indicates extensive hybridization (Arnold 1997:58-59).
Along the same lines, a rather intense debate has emerged in recent years regarding the extent of hybridization between wolves and coyote. Wayne (1995) and Wayne and Jenks (1991) claim that the red wolf is nothing more than a coyote-gray wolf hybrid. Wayne and Gittleman (1995) examined red wolf (Canis rufus), gray wolf (Canis lupus) and coyote (Canis latrans) nuclear DNA and mtDNA and found no genetic trait in the red wolf that was unique to that species. Red wolf mtDNA was found to be nearly identical with the mtDNA of coyotes examined in Louisiana (see also Lehman, et.al. 1991; Wayne et.al. 1991; Roy et.al. 1994). Nowak (1995) criticizes Wayne and Jenks' position and argues instead that the hybridization of the red wolf and coyote originated in Texas only 100 years ago and spread to the red wolf's former range. At the heart of the debate, however, is not whether hybridization has occurred among these species, but rather how long it has been taking place; the interfertility of wolves and coyote is more or less universally accepted (see Mayr 1996) (see Figure 2). Similar disagreements exist regarding how long and how extensively dogs and arctic wolves have interbred (see Wilson and Reeder 1993; Nowak 1995).9
Figure 2
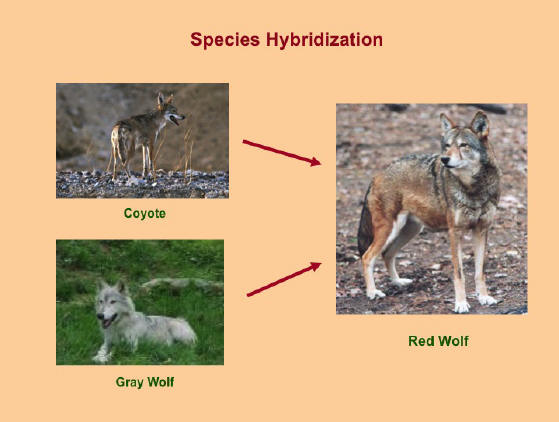
Gray Wolf-Coyote Species Hybridization
Thus, despite the persistence and pervasiveness of apparent species distinctions within local ecological communities, considerable interbreeding and exchange of genetic information clearly occurs among closely related species populations. It even appears as though many existing species originated as a result of hybridization in their evolutionary past. Indeed, Arnold and Meyer (2006:273) proposed that, due to the ubiquity of natural hybridization, the "web" rather than the "branching tree" provides the most appropriate metaphor for portraying phylogenic evolution.
Further Complications
Several additional phenomena illustrate the absence of a clear genetic definition of species, including circular overlap, asymmetrical sterility, sibling species, apomixis and self-fertilization.
Circular overlap occurs when "a chain of intergrading subspecies forms a loop or overlapping circle of which the terminal links have become sympatric without interbreeding, even though they are connected by a complete chain of intergrading or interbreeding populations" (Mayr 1963:507) (see Figure 3).
Figure 3
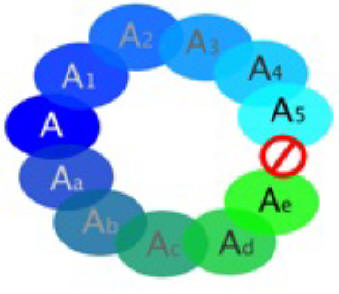
Circular Overlap
This process is illustrated by the California salamander (Ensatina eschscholtzii) (see Figure 4). This species, originally located in northern California, expanded southward from its ancestral territory along two different mountain ranges separated by the Central Valley of California. E. eschscholtzii now consists of seven subspecies, two of which exist sympatrically in southern California where they not only differ in appearance but also generally fail to interbreed (Irwin, Irwin and Price 2001:235).
Figure 4
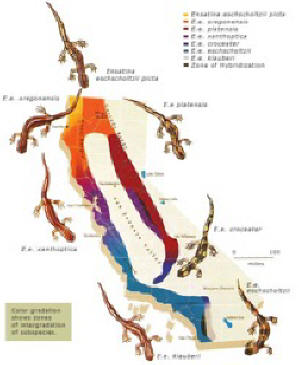
E. eschscholtzii Circular Overlap
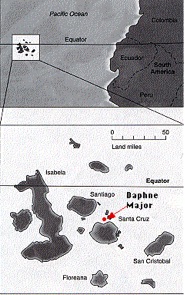
Species such as E. eschscholtzii, which exhibit circular overlap, are referred to as Ring Species. As with hybridization, the occurrence of ring species is relatively common, especially among birds and fish. (see Wiltenmuth and Nishikowa 1998; Irwin, Irwin and Price 2001; Bowen et.al. 2001; Irwin and Irwin 2002), and their existence has been recognized for more than a century (see Jordon 1905; Cain 1954). The large tree-finch (Camarhynchus psittacula) and the medium tree-finch (C. pauper) exist sympatrically on Floreana Island in the Galapagos without interbreeding. However, these two species, like E. eschscholtzii, are connected through a series of intermediate forms on the other nearby islands that can and do interbreed (see Futuyma 1998:155; see Figure 5).
Figure 5
Camarhynchus Circular Overlap
Similarly, two species of ringed plovers (Charadrius hiaticula and C. semipalmatus), which display a circumpolar distribution, coexist in the eastern region of Baffin Island without interbreeding. However, to the east and west of Baffin Island they are connected by a ring of interbreeding hiaticula and semipalmatus populations respectively (Bock 1959:98-100).
In all, some 23 taxa have been proposed as ring species, including 12 avian species (Irwin and Irwin 2002:598). Ring species status has also been proposed for amphibians (salamanders and frogs), rodents (deer mouse and house mouse), insects (bees, butterflies, tsetse fly and drosophila) and fish, as well as for several plant species. The varied interfertility associated with those local species populations experiencing circular overlap illustrates incipient speciation. Indeed, so important is circular overlap to the origin of species that Mayr (1942:180) called it the "perfect demonstration of speciation." However, for the very reasons that ring species illustrate the process of species diversification, they invalidate the notion that species status can be simply defined by the presence or absence of interbreeding, as the ability or inability to breed evolves over time.
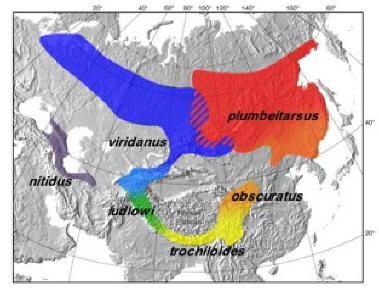
Ring species present a conundrum for systematists attempting to classify species: they simply don't fit into the existing species classification structure. Two sympatric populations can, on the one hand, be considered separate species where they coexist because they don't interbreed; yet, they can be defined as members of a single species because they are connected by a chain of fully interbreeding local populations. This has resulted in considerable disagreement over the classification of several ring species. For example, while some researchers view the California salamander (E. eschscholtzii) as a single species (cf. Stebbins 1949; Wake and Schneider 1998), Highton (1998) argues that E. eschscholtzii should be divided into eleven different species. Similarly, the ring of Greenish Warblers encircling the Tibetan Plateau (see Figure 6) is classified by some taxonomists as a single species, but as two distinct species by others (Irwin, Irwin and Price 2001:240). Likewise, local populations of the Arctic Gull (see Figure 7) are variously classified as different species or as sub-species, even though molecular markers show little differentiation between them (Irwin, Irwin and Price 2001:235).
10 However, as each of these examples clearly illustrates, the very attempt at classification --whether as one, two or more species-- obscures the most significant phenomenon from the perspective of evolution: the differential interbreeding and gene flow that occurs among local populations. It, thus, undermines a meaningful understanding of the fundamental role that ecological processes play in creating and maintaining locally distinct populations. It is for this reason that the term circular overlap is preferable to ring species; the former focuses on the dynamic process rather than on a static typological classification.
| Figure 6
Greenish Warbler Circular Overlap
(http://www.zoology.ubc.ca/~irwin/GreenishWarblers.html)
|
Figure 7
Arctic Gull Circular Overlap (Irwin, Irwin & Price 2001:235) |
Asymmetrical sterility occurs when different levels of fertility prevail, depending on which sexes are involved in the mating of individuals from two or more populations. Asymmetrical sterility is illustrated by the species pipiens (see Mayr 1963:42). Five varieties of this species exist in Europe: one in western Europe, one in northern Germany, one in southern Germany, and two along the Mediterranean. Males of the southern German variety experience full fertility when mated with females of the western European and northern German varieties. However, reciprocal crosses between females of the southern German variety and males of the western European and northern German varieties are sterile, with less than 1% interfertility. To further complicate the issue, American pipiens experience complete fertility when crossed with the northern German variety, but are more or less sterile when mated with the other four European varieties.
A similar situation occurs among butterfly species of the genus Heliconius in Central America. According to Jiggins et al. (2001), female offspring are fertile when males of the Panama species are crossed with females of the French Guiana species, achieving a hatch rate of .88 compared to hatch rates of .90 for conspecific French Guiana matings and .93 for conspecific Panama matings. However, the female offspring are sterile when males from French Guiana are crossed with females from Panama. These latter female offspring show no obvious abnormalities, except for the fact that their eggs are considerably smaller than normal. Significantly, male offspring are fertile whichever direction the crosses occur. A comparable breeding relationship exists between the French Guiana species and the species from Columbia; female offspring of Columbian males and French Guiana females are fully fertile, while female offspring of French Guiana males and Columbian females are sterile, with most of the Columbian females never even laying eggs (see also Naisbit et al. 2002).
Grula and Taylor (1980) discovered comparable examples of asymmetric female sterility between Colias eurytheme and C. philodice, two species of sulfur butterflies that inhabit much of North America. Although the two species display strong ethological isolation wherever they exist sympatrically, they retain a high degree of genetic compatibility. However, while F1 females from the cross of C. philodice females and C. eurytheme males exhibit normal fertility and fecundity and display heterosis (hybrid vigor), F1 females from the cross of C. eurytheme females with C. philodice males are normally unviable and almost always sterile.11
Defining species boundaries has been further complicated by the ubiquity of sibling species (cf. Knowlton 1993; Brockhouse and Tanguay 1996; Finlay 2002; Mathews et al. 2002; Crummett and Eernisse 2007). Sibling species are those species that differ little in physical appearance, but display quite distinct breeding habits, habitat preferences, and other defining characteristics (see Mayr 1963:33-58; 1976:509-514). A study of light flashes among fireflies of the genus Photuris, for example, resulted in the classification of 18 species where previously only 2 or 3 species had been recognized on the basis of physical appearance (Barber 1951). This reclassification led to the discovery of several additional characteristics associated with the variation in light flashes, including breeding season and habitat preference. Observing the mating songs of green lacewing crickets (Chrysoperla), Wells and Henry (1992a) concluded that three distinct species existed where only one had previously been defined in terms of morphology (see Futuyma 1998:458). Similarly, examining habitat differences Andersson (1990) proposed 16 species in the genus Maranta (prayer plants) in the American tropics where previous authors had recognized only 5.12
Apomixis further complicates a genetic definition of species (see Cronquist 1978:11; Grant 1981; Gornall 1997). Apomixis is a form of asexual reproduction in which plant embryos grow from egg cells without being fertilized by pollen, the male part of the plant. Parthenogenesis is one type of apomixis that occurs in animals. It is similar to self-pollination in that the same genotype is perpetuated in successive generations, resulting in highly individualistic genetic lines. Like self-pollination, it is also not fully obligate, meaning that a small amount of sexual reproduction does occur. Defining species status for apomictic plants can, therefore, not be based simply on interfertility; otherwise every distinct self-pollinating and apomictic genetic line would be defined as a distinct species. According to Gornall (1997:175), "In many cases, these lines are morphologically distinguishable and it would be possible to give each a name at species rank, thereby recognizing them as microspecies." Grant (1981) further states that such an approach could be defended in cases where incompatibility barriers exist between the microspecies, as they do for example among several species of Gilia in the G. inconspicua complex.13
Self-fertilizing plants, such as those of the genus Crataegus,14 further complicate the definition of species. Due to the lack of a clear genetic basis for defining species status and to the lack of clear standards for species determination, considerable variation exists in the delineation of self-fertilizing and apomictic species, with "splitters" proposing more and "lumpers" proposing fewer species. At one point, according to Camp (1942:368), over 1,100 species had already been identified in the genus "with the end apparently nowhere in sight." Different taxonomists have proposed wildly different numbers of Crataegus species in Florida alone, ranging from 11 to 72 (ibid.). Today, the number of species recognized within Crataegus is closer to 200, though the exact total is still undetermined (Phipps et al. 2003). However, some sources still identify over 1,000 species, with new species continuing to be listed.
Methodological Issues
Aside from the complications introduced by the existence of circular overlap, asymmetrical sterility, sibling species, apomixis, self-fertilization and the many other natural phenomena associated with the diversity of living organisms, methodological problems arise with any attempt to determine species boundaries. Several authors have noted the numerous practical difficulties of defining species boundaries using tests of interfertility (Sokal and Crovello 1970; Van Valen 1976; Cronquist 1978; Andersson 1990; Boxhorn 1995). Sokal and Crovello (1970), for example, emphasize the central role that morphological considerations, subjective criteria and arbitrary decisions play in various testing procedures, underscoring the lack of a clear genetic foundation in species classification. The very sampling procedures used to test interfertility, they argue, must be based on observable, phenetic (not genetic) characteristics. Interfertility also cannot always be determined by a clear and unambiguous decision. Individual researchers have relied on different criteria to determine interfertility among species populations. Some researchers require only penetrance or the production of a zygote to determine conspecificity, whereas other researchers necessitate the birth of a viable offspring or, more stringently, the reproduction of a fully fertile offspring that can transfer the newly recombined genetic information to the next generation. Furthermore, inasmuch as successful interbreeding varies quite widely during interspecies crosses (see Cronquist 1978:10),15 as it does in conspecific matings, researchers who may agree on which of the above criteria should be used to define interfertility may disagree on the proportion achieved within a particular level that is required to qualify as successful interbreeding (Sokal and Crovello 1970).16 Since there are no clearly agreed upon standards for establishing species status using tests of interfertility, the procedures used to determine what level to choose and what proportion of successful matings within a specific level constitutes interfertility are both arbitrary and subjective and vary considerably among researchers in the field.17 Confusion has even been added to classification procedures by the existence of marked sexual dimorphism in many species as, for example, between the mallard male and female. This has, at times, resulted in the classification of the different sexes of the same species as distinct species.
Boxhorn (1995) illustrates the fundamental methodological problem in determining species interfertility. Using the Wisconsin bluegill sunfish as an example, he demonstrates the practical impossibility of actually determining how many species of bluegill sunfish exist in Wisconsin. Boxhorn (1995:2-3) calculated that 499,500 separate crosses would need to be undertaken in order to determine whether the members of any single species located in 1,000 different lakes in Wisconsin could interbreed with members of other species. However, Boxhorn argues that the above number of crosses would need to be doubled to 999,000 in order to accommodate the problem of asymmetrical sterility. Furthermore, even if just three replications of each test were conducted, in order to compensate for the fact that interfertility varies among individual members of local species populations, the total number of tests needed would have to be increased to nearly 3 million, a highly impracticable number. However, he argues that even this number would not be sufficient to determine with any degree of confidence the extent of interfertility that exists among the various bluegill sunfish populations located in those 1,000 lakes, as well as with other closely related species populations. Inasmuch as Wisconsin contains over 16,000 lakes and ponds, Boxhorn claims that those 3 million tests would have to be increased to perhaps 48 million, and even then researchers would not fully know how many species of bluegill sunfish exist, since the bluegill sunfish also lives in the other nearby states. In other words, the number of crosses needed to adequately determine species boundaries in any single case study is likely to be astronomical, meaning that, for all intents and purposes, determining species status by tests of interfertility is for most cases highly improbable, if not impossible.
Defining species boundaries, then, is by no means clear and obvious, simply a matter of reporting the evident genetic discontinuity in nature. The generally held notion that species status can be clearly determined by whether or not a fertile offspring is produced does not in any way represent the species concept as it currently exists in the biological sciences. For the most part, fertility is not the principal criterion used to determine species status among local populations. Where considerations of fertility are employed, numerous complications arise which result in subjective decisions being made by individual investigators that necessarily introduce arbitrary criteria into species classification procedures. This reality of species classification severely undermines any notion of the precise genetic basis of species. In most cases, hybridization, circular overlap, asymmetrical sterility and other evolutionary processes, combined with subjective and arbitrary classification procedures and the impossible number of crosses needed to make clear determinations, demonstrates that most species boundaries are neither precise nor genetic.18 Given the fact that evolution occurs slowly over time and that nature does not divide into neat categories, the actual occurrence of well-circumscribed, clearly definable species is the exception rather than the rule (Mayr 1963:90; Sokal and Crovello 1970:148). To fully appreciate the indeterminacy of species as a biological phenomenon, it is important to understand the confusion and conflict associated with the species concept in contemporary biological research.
The Species Problem
In order to appreciate the arbitrary and constructed basis of the species concept, it is important to understand the controversy that exists among biologists regarding the very nature and existence of species. Several major competing species concepts exist, including: the Morphological / Phenetic Species Concept (MSC),19 the Biological Species Concept (BSC),20 the Ecological Species Concept (EcSC),21 and the Evolutionary / Phylogenetic Species Concept (EvSC / PSC)22 (see Dobzhansky 1935, 1937, 1970; Mayr 1942, 1963, 1970, 1996; Stebbins 1950; Simpson 1951, 1961; Van Valen 1976; Cronquist 1978; Cracraft 1983; Coyne et al. 1988; Andersson 1990; Grant 1992, 1994; Boxhorn 1995; Claridge et al. 1997; de Queiroz 2007). Several additional species concepts have also been proposed. Nearly 70 years ago, Camp and Gilly (1943) defined 12 different types of species based on their population structure and reproductive behavior. More recently, Templeton (1992) proposed the Cohesion Species Concept; Harlan and de Wet (1963) the notion of Compilospecies, Van Valen (1976:237) Mutualistic Species, Turesson (1922a, 1922b) and Turrill (1946) Ecospecies and Coenospecies; Paterson (1993) and Lambert (1995) the Species Recognition Concept (see also Coyne et al. 1988); Van Valen (1976) the concept of Multispecies; and Durden (1969) and Pigliucci (2003) Pigliucci and Kaplan (2006) the Family Resemblance Species Concept. Indeed, Hey (2001) tallied as many as 24 distinct species concepts (see also Mayden 1997).23 The fact that such a variety of species concepts exist and that the definition of species has generated heated debates clearly demonstrates a lack of agreement among professional biologists regarding the very nature and delineation of species.
Significantly, none of the species concepts that have been proposed over the past 100 plus years can be applied to all species, as each concept possesses serious limitations that preclude its universal application. Each species concept employs different criteria for defining species. While the MSC employs phenotypic criteria to classify species, the BSC defines species in terms of actual interbreeding in the wild (reproductive isolation). The EvSC, on the other hand, focuses on genetic relatedness, while the EcSC defines species in terms of their occupation of similar ecological niches. Inasmuch as the MSC classifies organisms as members of the same species based on phenotypic resemblance, many geographically separated organisms lacking any ongoing genetic interaction (and thus evolutionary continuity) would be classified as members of the same species using this concept.. At the same time, the BSC, which defines species as reproductively isolated genetic units, cannot be applied to asexual species without every individual organic line being classified as a separate species.
Furthermore, since each species concept applies different criteria to define what constitutes a species, the various species concepts lead inevitably to different species classifications. Thus, what is determined to be a distinct species using one species concept is not necessarily considered a separate species when employing a different species concept, and what might be considered a single species according to one species concept might be defined as several distinct species when using a different species concept. If, for example, a group of organisms becomes geographically separated (and thus reproductively isolated) from its parental population, such as oaks in Texas vs. oaks in Canada, the BSC defines the two populations as distinct species, even if they have not become ecologically or morphologically distinct. According to the MSC and the EcSC, however, these two populations are not defined as distinct species, even though they have become reproductively isolated, because they continue to resemble the parent population (MSC)24 or continue to occupy the same ecological niche (EcSC) (see Ereshefsky 2011:74-75). Similarly, the BSC, MSC and EcSC disagree as to whether asexual organisms form distinct species. According to the BSC they do not form species, whereas according to the MSC and the EcSC they are classified into species determined by their shared physical appearance and their shared ecological adaptation respectively, even if they are geographically isolated from one another (ibid.)
As a result, according to Ereshefsky (1998:105), the various species concepts "carve the tree of life in different ways. Many interbreeding species fail to be phylogenetic species, and many phylogenetic species fail to be interbreeding species." Freshwater fish represent an example of the former (see Rosen 1979), while asexual organisms constitute an example of the latter. Similarly, since Balsam Poplars and Cottonwoods have produced fertile offspring for 12 million years, despite the fact that they have remained ecologically distinct, the BSC recognizes the entire group as a single species, whereas the EcSC classifies them as two distinct species belonging to the same genus (Ereshefsky 1994:192).
There is also considerable variation in the degree to which the various species concepts agree or disagree with one another in the definition of species. Sometimes the BSC and the EcSC may recognize organisms as belonging to the same species, but not the EvSC. At other times the EvSC and the EcSC may agree, but not the BSC, or the BSC and EvSC may agree but not the EcSC. For example, while asexual organisms cannot form a species using the BSC, they can when employing the MSC, EvSC and EcSC. On the other hand, while the BSC allows the existence of ancestral species, the EvSC does not (see Ereshefsky 2010a:411-412).25 Similarly, a lineage of organisms lacking a common selective regime cannot form a species when using the EcSC, but can when using the MSC, EvSC or BSC (Ereshefsky 2011:72-73). Which species concept is employed is largely determined by the nature of the species being examined, by the research questions under investigation, and/or by the theoretical orientation of the researcher.
Finally, there is a critical problem shared by all of the existing species concepts; they have all been developed to delineate and classify eukaryotic organisms; none can be used to classify prokaryotic organisms, which constitute the majority of living organisms.26 Prokaryotes differ in important fundamental characteristics that render existing species concepts inapplicable to them (see Rosello-Mora and Amann 2001; Rosello-Mora 2003; Ereshefsky 2010b). Prokaryotes, for example, frequently exchange both homologous and non-homologous genes through a process known as Lateral Gene Transfer.
Lateral Gene Transfer is the movement of genes between distantly related organisms. Such gene transfers can occur without sexual reproduction, as when a virus inserts its genetic information into the genome of a host organism. Although heritable lateral gene transfer occurs between microorganisms and their eukaryotic hosts (Hotopp, et al. 2007), it is not a characteristic of eukaryotic evolution. By contrast, it occurs quite frequently among prokaryotes and has been of evolutionary significance in this domain. Furthermore, the transfer of non-homologous genes occurs not only between organisms in distinct families, but also between organisms in completely different kingdoms. Such relatively open gene pools render the application of the BSC, with its emphasis on reproductive isolation as the defining species criteria, completely inapplicable to prokaryotic organisms. Similarly, given the relative frequency of non-homologous gene transfers from a variety of sources, different parts of a prokaryotic organism's genome could display distinct evolutionary histories, depending on which genes were chosen for phylogenetic analysis. This seriously undermines the application of the Phylogenetic Species Concept to prokaryotic organisms.
Because no single species concept can be applied to all species, or for that matter even to all eukaryotic species or to all prokaryotic species,27 most biologists have adopted a pluralistic view of species in which specific species concepts are seen as able to delineate one aspect of some species, but incapable of defining species as an entity in nature. The growing acceptance of species pluralism has led an increasing number of biologists, systematists and philosophers to question the very existence of species as a "real" natural unit and to call for the elimination of the species concept (see Ereshefsky 1998; 2002; 2010a, 2010b, 2011; Mishler 1999, 2003; Hendry, et al. 2000; Plejel and Rouse 2000a, 2000b; Fisher 2006).28 Of critical importance is the fact that the inconsistency displayed by groups of organisms united under the species category does not arise because a few species fail to possess the biological features found in most other species, but rather because the lack of coherence occurs throughout the organic world. That is, the problems associated with defining species are not the result of a few outliers, but rather with the majority of species. In other words, "the disunity of the species category is due to major discrepancies in the biological world, not a few exceptions" (Ereshefsky 1998:113). The problem, therefore, is clearly with the nature of the species concept itself and raises serious questions regarding its continued application.29
This was, in fact, the central point that Darwin made more than a century ago. While Darwin accepted those local species taxa defined by "competent naturalists" as real (Ereshefsky 2010a), he rejected the existence of the broader species category and characterized various attempts to define species as "laughable."30 In the Origin of Species, Darwin (1859 [1964]:52) stated that "I look at the term species as one arbitrarily given for the sake of convenience to a set of individuals closely resembling each other, and it does not essentially differ from the term variety." Indeed, in Chapter 8 of Origins, in which he discusses hybridization, Darwin (1859[1964]:248) states that "neither sterility nor fertility affords a clear distinction between species and varieties." In summing up the chapter, he concludes that "the facts briefly given in this chapter do not seem to be opposed to, but even rather to support the view, that there is no fundamental distinction between species and varieties" (ibid.:278). Hence, despite the title of his book, Darwin did not even once use the term speciation. He rejected any distinction between the processes leading to the evolution of those groups of organisms given the more significant label "species" and those given the lesser label "varieties."31
Species Are Not Evolutionary Units
The species has generally been viewed as the basic evolutionary unit, with the larger taxa representing the hierarchy into which all species can be placed (see Mayr 1942, 1963, 1970,1996; Wiley 1981; Ghiselin 1974, 1987; Eldredge and Cracraft 1980). This notion is largely based on the belief that species represents the basic reproductive/evolutionary unit, whereas the various larger taxa constitute the organization of those basic units into a nested hierarchy. However, species are no more basic reproductive/evolutionary units than are any of the other categories created to organize the diversity of living organisms. As already indicated, the basic reproductive unit among self-fertilizing plants is the individual organism. Introgression, hybridization and horizontal gene transfers further undermine the notion that species are basic reproductive units (see Ereshefsky 1991, 2010a, 2010b). Certainly, neither ecological, phenetic nor phylogenetic species would be defined as reproductive units, since the criteria used by these concepts to define species is not based on reproduction. Furthermore, since most species are made up of local populations that rarely, if ever, exchange genetic information, species cannot be considered reproductive units (see Donoghue 1985; Templeton 1992; Ereshefsky 2010a). It is the local breeding population adapting to local ecological conditions that constitutes the basic reproductive-evolutionary unit (see Mishler and Donoghue 1982; Ereshefsky 1991). Inasmuch as species are human taxonomic constructs (see Hey 2001), they are no more basic evolutionary units than are any of the other taxa within the Linnaean hierarchy. Species, like all other taxa, are genealogical-historical entities created by adaptive processes operating on local populations (Mishler 1999; Ereshefsky 1991, 1994, 1998, 2010a). For that reason, all taxa may be considered epiphenomena created to varying degrees by the cumulative history of past adaptations among local populations. Ehrlich and Raven (1969) similarly argue that taxa larger than the local population are not meaningful genetic units (i.e., breeding populations), but rather represent merely "phenetic clusters", that is, populations that share certain physical or behavioral characteristics, but that do not function as reproductive units. They attribute retained similarities among separated populations of the same species to the independent effect of similar selective pressures rather than to continued gene flow and note that strong selective pressures can act as a powerful evolutionary force even in the face of continuous and counteracting gene flow. Sokal and Crovello (1970;148) agree with this position and add that "nothing is to be gained by the additional abstraction to the species level, but much is lost, namely accuracy, for no two localized populations are alike."
Species status among local populations must, therefore, be viewed as variable and a function of local selective conditions, rather than as a reality qualitatively defined by specific a priori species-specific characteristics. Indeed, as with many of the attributes which function to distinguish and reproductively separate two closely related populations, the very degree of interfertility between two closely distributed species may vary from one local community to another, as the very process of speciation implies. Population biologists and ecologists have, therefore, concerned themselves with understanding the mechanisms and processes that produce changes in species boundaries (species differentiation) among local populations and the relation that such speciation and species replacement have to the evolution of encompassing ecological communities (see Mayr 1963, 1976; Levins 1968; Margalef 1968; Brookhaven National Laboratory 1969; Cody and Diamond 1976).
The Essentialist Nature of the Species Concept
The underlying problem with the species concept is that it constitutes an essentialist, Aristotelian concept developed within a creationist worldview. The Linnaean System of nomenclature, which is still used to classify species and to provide the framework within which species are perceived, originated more than a century before Darwin's Origin of Species and makes no reference or accommodation to evolution (Pleijel and Rouse 2000b:627). Central to the Linnaean essentialist conceptualization of species is the belief that species are distinct entities that are: (1) stable over time; and (2) possess a central essence common to all of its individual members. However, despite the rejection of its essentialist assumptions, the Linnaean hierarchy is still used in contemporary biology. It is the essentialist conceptualization of species associated with the Linnaean classification system that underlies the notion that species are: (1) "real", (2) clearly definable, and (3) the basic units of evolution. This has created a fundamental conundrum with regard to the conceptualization and classification of species within the natural order. As Ereshefsky (1994:199) notes, "The history of biology has left us with a hierarchy of categories without an ontological foundation."32 .
Within the Linnaean system, individual organisms can belong to only one species (Ereshefsky 1998:109-110). Consequently, the classification system and the conceptualization of species that it presents cannot accommodate either the contemporary pluralist conceptualization of species, in which more than one "correct" taxonomy exists, or the ubiquity of hybrid species (Ereshefsky 1994:193-195). The Linnaean system of classification also cannot accommodate Circular Overlap, Asymmetrical Sterility, and the many other phenomena that complicate the clear definition of species. Finally, since the decision regarding whether a group of organisms is recognized as a distinct species or not is dichotomous (i.e., a yes or no proposition), the Linnaean system and its essentialist view of species is incapable of expressing both the complexity and the subtlety accompanying the diversity of living organisms within a theoretically integrated evolutionary framework (Hendry, et al. 2000:72).
33
The many problems associated with the species concept, especially as it is presented in the Linnaean hierarchy, has led an increasing number of researchers, beginning with Darwin, to call for its elimination on the grounds that the concept is both irrelevant and obstructionist in explaining the origin and diversity of life. Fundamental to the use of any concept in science is that it has both predictive and explanatory value, as well being capable of clear operational definition. The species category possesses none of these attributes. Developments in biological evolutionary theory have rendered the species concept increasingly problematic. Concepts in science are theoretically constructed; consequently, as theory changes so also do the central concepts employed in any discipline. Like the concept of phlogiston in physics, entelechies in biology and race in anthropology, the concept of species has outlived its usefulness. Just as phlogiston was eventually abandoned in physics and race in anthropology, many have argued that so must the species concept be abandoned in biology as an outdated concept with underlying essentialist assumptions incompatible with modern evolutionary theory. Several researchers have proposed alternative concepts, including "biospecies" and "ecospecies" (Grant 1981:36), "phylospecies" (Ereshefsky 1998:117-118), "evolutionary units" (Hey 2001:328), and "Least Inclusive Taxonomic Units." (Pleigel and Rouse 2000a, 2000b).34
The numerous problems associated with the species concept have led to the development of a system of numerical taxonomy that makes no reference to species names, but rather organizes the various taxa exclusively in terms of quantitative similarities associated with various phenetic and genetic characteristics (Heywood 1967; Hendry et al. 2000). Such a system represents a non-essentialist and non-typological approach to organizing biological diversity that is more compatible with evolutionary theory. According to Hendry et al. (2000:74), "Studies of 'speciation' could be recast as studies of reproductive isolation, and of genetic and phenotypic divergence (e.g., how much isolation or divergence over how long a period)." Such an approach would allow a more subtle and realistic understanding of the evolution of biological diversity unencumbered by the limitations imposed by attempts to fit this variation and the evolutionary processes that have produced it into the straightjacket imposed by the reified species concept.
The position argued here is that species do not constitute a concrete phenomenon, but rather comprise a conceptual unit within a systemic view of nature whose individual manifestations are continually in a state of flux, that is, in a constant process of either becoming, maintaining or disappearing. Indeed, current estimates are that about 99% of the species that have ever existed on earth are now extinct. Because of their fluidity, the precise description of species is difficult, if not impossible, to determine in the real world. To claim that species are real is to reify and essentialize as "things" what are, in fact, transient components of a dynamic and evolving system whose individual parameters are both highly variable and capable of shifting rather abruptly, and in a diverse variety of ways in response to changing environmental conditions.
It is for this reason, for example, that the concept circular overlap is preferable to that of ring species. While the former focuses on the dynamic process, which is open to continuous flux and variation, the latter focuses instead on the "things" that the process creates at an arbitrary point in time from the perspective of evolution. Moreover, in its focus on the results rather than on the process, the concept of ring species transforms multiple independent local populations into a single encompassing entity. The concept of ring species represents an essentialist, typological definition of species, with the ring species being just another "type" of species. Circular overlap, on the other hand, does not reify the existence of an entity (species), but focuses only on a process that is at once highly variable and potentially reversible. It implies only that evolution is taking place, recognizing that there is not necessarily an end product, nor an entity, which exists independent of that process.
Many of the problems associated with efforts to define species ultimately lie in the attempt to impose a static or typological classification scheme on what are inherently variable, evolving phenomena. Some of the problems also result from the failure to distinguish between species as a taxon within a hierarchical classification scheme and species as populations in local ecosystems. In the hierarchy of categories (taxa), species are but one taxon used to reconstruct phylogenic evolution. However, the species taxon should not be seen as more real than any other category in the hierarchy. To a certain extent, species (as well as the other taxa) only appear real due to the widespread existence of reproductive isolation, as well as to the fact that many of the intermediate forms which would blur the boundaries among contemporary species are currently extinct. Regarding local populations, species status must be viewed as a variable, something rarely achieved as an absolute existence and ultimately dependent upon local selective conditions. Within this context, those characteristics that distinguish one local species population from another do not exist a priori, but rather are consequences of the underlying ecological conditions that made their difference adaptive. Moreover, the degree of difference may vary substantially among the same populations inhabiting different ecosystems.
Indeed, the very concept of evolution presupposes more or less blurred species boundaries.35 At the heart of the Darwinian Revolution (see Mayr 1972) is the notion that species are more or less distinguishable taxonomic units, constantly changing in response to the selective pressures imposed upon local populations.36 As indicated by the quote introducing this paper, Darwin himself viewed species as subjective categories without clearly definable boundaries, not fundamentally different than varieties.37 Darwin rather clearly expressed his view of attempts to define species in a letter he sent to Joseph Hooker, dated December 24, 1856. Given that contemporary attempts to define species have been no more successful than those of Darwin's day, it would seem that his judgment has stood the test of time.
"It is really laughable to see what different ideas are prominent in various naturalists' minds, when they speak of "species"; in some, resemblance is everything and descent of little weight, in some, resemblance seems to go for nothing, and Creation the reigning idea, in some, sterility an unfailing test, with others it is not worth a farthing. It all comes, I believe, from trying to define the indefinable."
--Charles Darwin (1856)
* * * * *
NOTES
1.
Reproductive isolation is phrased in terms of populations, not individuals. Just as individuals of different species may be capable of interbreeding at the same time that their respective populations remain reproductively isolated, so also might two individuals of the same species be unable to interbreed (e.g., two males, or a Great Dane and the Chihuahua), yet be accepted as members of the same species (Futuyma 1998:451).2. Karpechenko (1927, 1928), a Russian cytologist, even successfully crossed the radish (Raphanus sativus) with the cabbage (Brassica olercia) and produced a viable though sterile hybrid, even though the two plants are members of completely different genera (Boxhorn 1995:6).
3. Iris brevicaulis and Iris fulva have very different primary pollinators --bumblebees and hummingbirds respectively-- which limits their interbreeding in the wild. However, when crossbred in the laboratory, they produce a rich variety of flowers.
4. Knoblock (1972) identified 23,675 instances of hybridization among different species and genera of angiosperms. According to Arnold (1992:251), natural hybridization also occurs among different species of eucalyptus.
5.
While the common belief is that the offspring of interspecific matings are sterile, such as the belief that the mating of a horse and donkey results in a sterile mule, quite the contrary is true. Not only are fertile mules occasionally produced (c.f. Fan et al. 1992), but fertile ligers (Lion-tiger crosses) have also been produced.. In fact, the offspring of natural hybridization are quite frequently fertile.6.
Mayr has been quite consistent and adamant in his opposition to the significance of hybridization:"The total weight of available evidence contradicts the assumption that hybridization plays a major evolutionary role among higher animals." (Mayr 1963:133).
Mayr's opposition to the significance of hybridization stems from his view that species represent well-integrated genetic systems whose viability would be undermined by hybridization.
"hybrids between species, particularly in animals, are almost always of inferior viability and more extreme hybrids are usually even sterile. 'Almost always' means that there are species interpreted to be the result of hybridization, particularly among plants, but except for the special case of allopolyploidy, such cases are rare." (Mayr 1996:264)
7.
Grant and Grant (1994:313) view speciation through hybridization as a form of founder effect that is consistent with the type of rapid evolutionary changes proposed by Eldridge and Gould (1972) in their Punctuated Equilibria model of evolution.8.
This represents, in some ways, a new, more theoretically sound, take on the older notion of "hopeful monsters," the concept proposed by Goldschmidt (1940) to characterize the theory that sudden jumps in evolution are necessary to explain how new biological species arise.9.
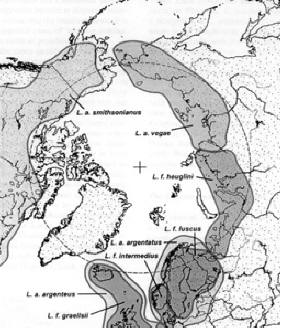
Debates over canid hybridization have largely been precipitated by the implications they raise regarding species conservation. Hybrid species may not be acceptable candidates for conservation. However, regardless of their political implications, or of the timing of gray wolf-coyote and/or arctic wolf-dog hybridization, it is clear that these various species have not been defined by their inability to interbreed and produce viable and fertile offspring.10. The circumpolar group of Larus gull species has frequently been cited as a classic example of ring species. The range of these gulls forms a ring around the North Pole. The Herring Gull, which lives primarily in Great Britain, can hybridize with the American Herring Gull (living in North America), which can also interbreed with the Vega or East Siberian Herring Gull, the western subspecies of which, Birula's Gull, can hybridize with Heuglin's gull, which in turn can interbreed with the Siberian Lesser Black-backed Gull (all four of these live across the north of Siberia). The last is the eastern representative of the Lesser Black-backed Gulls back in northwestern Europe, including Great Britain. However, the Lesser Black-backed Gulls and Herring Gull are sufficiently different that they cannot interbreed; thus the group of gulls forms a continuum except in Europe where the two lineages meet. However, a recent genetic study has shown that this example is far more complicated than presented here, and probably does not constitute a true ring species (Liebers et al., 2004). (see Larus)
11.
F1 refers to females of the first generation. Two criteria were used to determine fertility: egg color and larval survivorship. Eggs that turned pink within 24 hours were judged to be fertile, whereas those that remained white were determined to be sterile. In the absence of data on egg color, the appearance of second instar larvae was defined as a positive indication of fertility. A mated pair was considered normally fertile if more than 90% of the eggs produced turned pink or if second instar larval survivorship was greater than 30%, the normal larval mortality rate of the parent species raised under similar conditions. Whereas F1 females from the cross of C. philodice females and C. eurytheme males (Type 1) exhibited levels of fecundity and fertility comparable to that of the parent species, 17 out of 23 of the F1 females from the cross of C. eurytheme females with C. philodice males (Type 2) that mated and were tested for oviposition laid no eggs (Grula and Taylor 1980:675). Similarly, while Type 1 F1 females exhibited heterosis, including stronger flight and greater longevity than individuals in either parental species (over a month compared to two weeks), Type 2 F1 females were weak fliers and had lifespans slightly shorter than either parent species (ibid: 680). Equally significant is the fact that these dichotomies were also present in F2 females and in offspring produced when Type 1 and Type 2 females were backcrossed with members of their parental species.
12.
Grant (1992:311) argues that Andersson defines some populations as species that could just as easily be considered races of the same species, given the criteria he applies. Grant's criticism of Andersson reinforces the notion discussed previously that the definition of species involves a fair degree of arbitrariness and is a function of the individual researcher doing the defining. He later claims that the array of recognized population systems has outgrown the existing system of nomenclature. In describing the various categories employed, he includes, contiguous geographical races, disjunct geographical races, ecological races, allopatric semi-species, sympatric semi-species, allopatric species, sympatric species, superspecies and syngameons. As will be seen later, species definitions are fundamentally determined by which species concept is being employed.
13.
Gilia is a genus of between 25 and 50 species of flowering plants native to temperate and tropical regions of the Americas. Gilia inconspicua is a species of flowering plant known by the common name "shy gilia," which is native to the western United States.
14.
Crataegus, a well-known herb commonly called hawthorn or thornapple, is a large genus of shrubs and trees in the rose family, Rosaceae.
15.
"Members of different local populations may show a complicated and irregular pattern of interfertility and intersterility. Local population a may be fertile with b, d and f, but not with c and e." (Cronquist 1978:10). By the same token, population b, which is interfertile with a, may also be interfertile with c and e, but not with d and f.
16.
Recourse to arbitrary decisions is not uncommon in the application of methods used to define species.. In their description of how the Phylo-phenetic Species Concept is applied, Rossello-Mora and Amann (2001:59-60) indicate that a particular genetic marker is employed to construct phylogenetic trees. Then DNA:DNA hybridization is used to identify groups that have "a high degree of genomic similarity" (ibid.). A 70% degree of similarity serves as their threshold for identifying species. This number is chosen for practical, not theoretical considerations: phenotypic and genotypic similarities tend to agree with one another at 70% or higher. However, if there is a disagreement between the phylogenetic analysis and the 70% hybridization threshold, the threshold is simply reduced. At the same time, a phenetic analysis of as many characters as possible is employed, but again there is no theoretical basis for which characteristics are used (Ereshefsky 2010b:558). In the end, subjective and arbitrary decisions are made regarding both which characters are analyzed and what hybridization levels are applied to determine species status. In no case does an objective theory of evolution guide their analysis.
17.
The arbitrariness associated with establishing species status is illustrated by several examples. Merriam (1918) recognized 78 species of brown bears in North America, which are currently considered a single species (Cronquist 1978:4-5). Similarly, Phacekia ranunculacae is divided into distinct geographically separated populations with different numbers of chromosomes (6 vs. 14) and have resisted all attempts to hybridize them (Cronquist 1978:14), Chuang and Constance (1977) have unequivocally defined them as conspecifics on the basis of their strong morphological similarity.
18.
Species definition is even influenced by national divisions. Heywood (1967:32) points out that the division of Europe into a large number of independent states, "each with its own traditions and schools of taxonomy," results in similar plants being classified differently. Great Britain, for example, abandoned the use of the sub-species category, regarding it as a "Continental peculiarity" (ibid.).
19.
The Morphological Species Concept (MSC) (also referred to as the Phenetic, Typological and Essentialist species concept) is based on a consideration of the physical and behavioral characteristics associated with a species population (see Cronquist 1978). The MSC predominated historically and continues to be applied by many biologists and naturalists, especially botanists. For Cronquist (1978:15-16), "species are the smallest groups that are consistently and persistently distinct, and distinguishable by ordinary means." A single specimen (the type specimen) provides the basis for defining a species. The emphasis is, thus, on the type rather than on the population. The MSC is highly problematic. First of all, morphological characters are not reliable indicators of species difference. As demonstrated by the existence of sibling species, Significant behavioral, reproductive and ecological attributes of species populations may not vary consistently with morphology, meaning that there is no inherent relationship between morphological and genetic differences. Similarly, where geographic variants of the same species display distinct physical or behavioral characteristics, they would potentially be viewed and classified as different species, regardless of ecological or genetic considerations. In addition, the use of different definitional criteria could easily result in different species classifications. The MSC also presents a problem when applied to self-fertile or apomictic species, where phenotypic traits can vary quite widely, as was shown in the previous discussion of hawthorns. Furthermore, a strict application of the MSC cannot easily accommodate species that are sexually dimorphic or that have distinct life stages, such as caterpillars. Cronquist (1978:16) fully accepts the problems inherent in the MSC: "The definition . . . is so generalized that it leaves a great deal of room for individual interpretation and difference of opinion. In a sense, it merely sets the limits within which reasonable taxonomists may disagree. I will answer that such is the nature of the world." Of greatest significance, however, is the fact that the MSC represents an essentialist (typological) view of species that does not approach species populations as inherently variable, dynamic and evolving systems. Because it is based on typological thinking rather than on population thinking, the MSC cannot be effectively integrated into either ecological or evolutionary theory. It cannot, therefore, be used to explain the origin and differentiation of species.20.
According to the Biological Species Concept (BSC), species are defined by "reproductive isolation," that is, by whether or not interbreeding normally occurs between members of two different populations in the wild. Such a definition renders the BSC completely inapplicable to both asexually reproducing organisms, and to allopatric (related but separated) populations, such as oak trees in Texas and Canada, or squirrels in Pennsylvania and Georgia. Mayr (1942, 1963, 1996) consistently argued that only sexually reproducing organisms comprise species, as have Ghiselin (1974), Bock (1992, 2004), Eldredge (1993) and others. Mayr (1942:121-122), the foremost proponent of the BSC, stated quite explicitly that "The known number of cases in which the above species definition may be inapplicable is very small, and there seems to be no reason at the present time for watering our species concept to include these exceptions." Mayr was clearly incorrect in his assessment, as the overwhelming majority of living organisms by any measure (biomass, number of species, etc.) reproduce asexually. In addition, the number of related populations (such as the oaks and squirrels mentioned above) living in separate habitats where interbreeding cannot occur is incalculable (see Groves 2001). There is also no evidence whatsoever of a sudden hiatus in reproductive isolation at the species level. Rather the degree of reproductive isolation increases gradually throughout the continuum of population differentiation (see Mallet 2008). Finally, the BSC cannot be applied at all to fossil species, as there is no way to assess whether such populations interbred or not.21.
Proponents of the Ecological Species Concept (EcSC) define species as "a lineage (or closely related set of lineages) which occupies an adaptive zone minimally different from that of any other lineage in its range and which evolves separately from all lineages outside its range" (Van Valen 1976:233). Van Valen uses the term "adaptive zone" to represent an ecological niche. Both Van Valen (1976) and Andersson (1990), the two principal proponents of the EcSC, distinguish the EcSC from the BSC and argue that, unlike the BSC, the EcSC gives full weight to the ecological factors involved in species differentiation. From the perspective of the EcSC, reproduction plays a far less important role in defining species than it does for the BSC. Rather, "species are maintained for the most part ecologically, not reproductively" (Van Valen 1976:235; see also Ehrlich and Raven 1969). Van Valen uses the genus Quercus (oaks) to illustrate his point. According to Van Valen (1976:235), "Quercus marcrocarpa in Quebec exchanges many more genes with local Q. bicolor than it does with Q. macrocarpa in Texas." It is, therefore, ecological selection, despite their geographic separation and lack of reproduction that maintains the single-species status of geographically separated populations of Q macrocarpa and that keeps Quercus marcrocarpa and Q. bicolor largely discrete in Quebec. Grant (1992) disagrees with this argument and states quite emphatically that all of the major proponents of the BSC have emphasized the role that ecological differentiation plays in the process of reproductive isolation between species. Grant further argues that the application of the EcSC is undermined by the lack of any rigorous, a priori definition of the ecological niche.22.
According to the Evolutionary / Phylogenetic Species Concept (EvSC / PSC), species constitute the smallest diagnosable cluster of organisms descended from a common ancestor and possessing a combination of defining or derived traits (see Cracraft 1983; Baum and Donoghue 1995). Unlike the BSC, the PSC can be applied to both asexual and allopatric populations. Also, by treating sub-species as potential species, the PSC reinforces Darwin's argument that varieties and species are quantitatively, not qualitatively, different. Consequently, The PSC has largely replaced the BSC as the predominant species concept used today. However, the PSC has its own problems. A major difficulty is determining which criteria qualify to define individual organisms as "diagnostically distinct." Researchers have defined species based on both genetic distance and morphological difference. However, neither genetic distances nor morphological traits can be considered discrete characters. There is also no specific percentage difference in genetic material that consistently reflects species-level differentiation. Furthermore, since genealogical discordance can exist at different locations on the genome, principally as a result of hybridization and introgression, different parts of the genome may display distinct genetic histories (see Tattersall 2007). This would result in different species delineations, depending on which genetic markers were used for definitional purposes. The PSC is also vulnerable to arbitrary species identifications, because some threshold of difference must still be chosen, which will vary among individual researchers. Which traits and how many traits (genetic or morphological) are necessary to define different species also remains an open question. The result is a lack of clear guidelines as to where in the taxonomic hierarchy individual populations should be divided into distinct species. Using morphological characters also raises not only the issue of defining such characters, but also of choosing which characters are diagnostically important. Since no direct relationship exists between the degree of morphological difference and either genetic distance or the ability to interbreed, using morphological variation as a criteria for defining species difference is both illogical and theoretically invalid. Chimpanzees, for example, share more genes in common with humans than they do with gorillas, despite the the fact that they are clearly more similar morphologically to the latter. This method is also undermined by the existence of sibling species, which are similar morphologically but different genetically. The increased application of the PSC has also resulted in "taxonomic inflation." There has been a dramatic increase in the number of recognized species, especially primate species, as a result of the shift in popularity from the BSC to the PSC (see Alroy 2002, 2003; Harris and Froufe 2005; Isaac et al. 2004; Knapp et al. 2005; Tattersall 2007). The number of primates species defined in the Smithsonian Institution's Mammal Species of the World increased from 181 in 1982 to 233 in 1993 and 356 in 2001, a 100% increase in just 20 years.